[From Bruce Abbott (2013.12.29.2225 EST)]
Rick Marken (2013.12.29.1600)–
Bruce Abbott (2013.28.1745 EST)
BA: I’ve been doing some on-line reading of the literature on the equilibrium point (EP) model and can see why Latash et al. would not be aware of any connection of their model with PCT.
RM: It’s not that they are “not aware” of a connection; they explicitly reject a connection because they reject control theory.
BA: In fact, I’ve found it difficult to understand their model, given the descriptions I’ve read of it. Thus far I have not seen a discussion of how the “commands” of the system are derived; it could very well be an open-loop system at that level, or maybe not.
RM: I’m pretty sure it’s an open loop model. Their “commands” are not like reference signals because they not specifications for input. Rather, they are commands for output – they cause the tensions in the muscles (outputs) that determine the equilibrium point of the limbs to which the muscles are attached.
BA: It’s like presenting a PCT diagram of a first-order control system without indicating what determines the reference signal values – they could be determined by the output of an open-loop or closed-loop system.
RM: I think it’s like presenting a PCT diagram of a first order control system without the first order perception and comparator. In other words, it’s nothing like a PCT diagram; it’s a diagram of a motor output system and I can only assume that they persist in going with this approach because they want to explain control in lineal causal terms. The relationship of Latish et at to PCT is one of complete opposition.
BA: You might be able to gain some clarity about the EP model by reading at least a portion of a paper by Perrier, Ostry, and Laboissiere (1996, which is available online at http://www.psych.mcgill.ca/labs/mcl/pdf/jshr96final.pdf.
BA: These authors discuss control jaw movements rather than arm movements, but they do illustrate how the EP model resists the effect of loads. They offer a simplified description for a single (closer) muscle first, then go on to treat antagonistic muscles. I’ve reproduced their Figure 1 below, which refers to the single-muscle case:
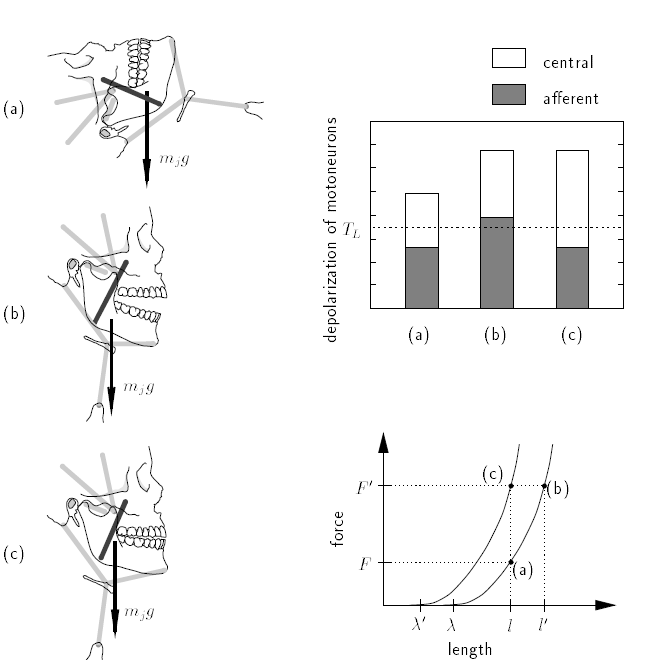
BA: In (a) the face is horizontal so the jaw is unloaded and the individual is holding the jaw slightly open. The bar graph shows the contributions of central input and afferent input (from muscle stretch receptors) to motor neuron depolarization, which determines the strength of muscle contraction. The line graph below the bar graph indicates the equilibrium-point functions, the rightmost being the curve determined by the current central input (λ). (a) on that curve refers to the equilibrium point established under the condition shown in the (a) figure of the skull and jaw.
BA: In Figure 1b, the face is not vertical and a downward force is being exerted on the lower jaw by gravity. With no change in the central input, the jaw is pulled open, stretching the muscle that closes the jaw, which increases the input from the stretch receptor. There is more total depolarization as shown in the bar graph at (b), so muscle force increases, thus resisting further opening of the jaw. Because the central input is unchanged, we are still on the same EP curve but now at (b) as shown in the lower graph.
BA: In Figure 1c, central input changes to establish a new EP curve (λ’ in the lower graph) As the jaw closes, the muscle shortens, so afferent input from the muscle spindles decreases to the same level seen in Figure 1a, but there is now more central input, so muscle length decreases while maintaining the same level of force and continuing to balance the force generated against the force due to load.
BA: You can get a more detailed description by reading the paper, especially the section labeled “Basic Control Mechanisms.”
Bruce