[From Bruce Abbott
(2009.05.15.1720 EDT)]
Bill Powers (2009.05.15.0203 MDT)–
Bruce Abbott (2009.05.12.1140 EDT)
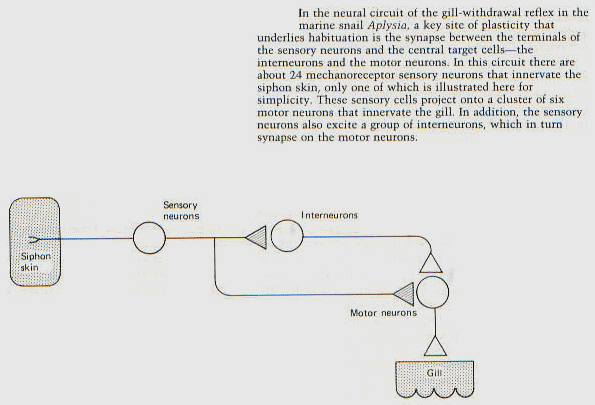
BP: Unfortunately, a diagram like that doesn’t tell us what the circuit does
(as I’m sure you know). If we don’t know the functions performed by each
neuron, we can’t tell whether this is a differentiator, a two-level control
system, or any of a large number of other possibilities. Are there any more
connections that were just not drawn?
BA: I’ve wondered about the role of the interneuron in
that diagram. My best guess is that it prolongs or intensifies the
contraction of the mantle muscles. (I haven’t yet read enough of this
literature to know what Kandel and colleagues have determined its function to
be within this circuit.) All the synapses in the diagram are excitatory, by the
way.
BP: The attached gives a little more information. I haven’t located the Figure
yet. It seems that the interneuron ONLY affects the amount of neurotransmitter
produced by impulses in the other axon – it does not synapse directly with the
motor neuron. So it appears to be purely a gain control.
Yes.
BP: One thing is pretty clear: this is not a
“learning” phenomenon. Associating it with Hebbian learning is just a
mistake, it seems to me. This system always does the same thing; it doesn’t
acquire any new way of reacting to inputs. It habituates and dishabituates over
and over, the same way every time, without learning a thing.
It’s
not Hebbian learning, and no one claims that it is. Evidence for Hebbian
learning has come from further studies of aplysia using a classical conditioning
paradigm, in which two stimuli have to be associated. Habituation and
sensitization do not involve an association and for this reason are referred to
as forms of “nonassociative” learning.
Whether
to call habituation and sensitization “learning” at all depends, I
suppose, on how you define learning. They do involve a change in behavior as a
result of experience, meeting the traditional definition of learning. In
short-term habituation and sensitization, the only thing that changes is what
might be viewed as the “weight” of the synaptic connections.
Connections are neither formed nor destroyed. In the long-term forms of these phenomena,
however, the number of synapses between the two neurons involved changes. Not
observed are synapses forming between neurons that formerly did not have such
synapses. So the “wiring diagram” still remains essentially
unchanged.
In
the “arm” demonstration that accompanies LCS III, reorganization
acts in the same way on existing connections, increasing the weights of some
and decreasing the weights on others. If habituation and sensitization do not
qualify as learning on the grounds that the wiring diagram doesn’t
change, then it seems to me that the form of reorganization implemented in the arm
demo doesn’t quality as learning, on the same grounds. My view is that
learning is involved in both cases.
BP: Gently squirting water onto the mantle
with a WaterPik resulted in withdrawal of the gill and the siphon. Repeated
squirts lead to less and less response, and eventually none. The interpretation
is that Aplysia “learned to ‘ignore’ the stimulus… because it was not
harmful.” I doubt that statement rather thoroughly.
BP: Apparently the normal case is for an
occasional stimulation of the mantle to cause withdrawal of the gill and
siphon. This says that the gain is being maintained at some fairly high level
by activity in the interneuron that raises the gain. Since that interneuron is
activated by sensors in the tail, there must be occasional stimulation of the
tail under these same normal circumstances. It is not unreasonable to assume
that when Aplysia is moving about, the tail is being stimulated.
BP: When the mantle alone is stimulated, but
not the tail, the gain begins to decline gradually because there is no signal
in the interneuron to maintain the gain at a high level. This explains the
habituation. If the tail is then stimulated, the gain is increased and the
habituation is abolished: the normal response to a combined tail and mantle
stimulation returns. The gain control seems to respond rapidly to a brief
signal from the tail, raising the gain, with the gain dropping slowly after the
signal. So only a low rate of stimulation of the tail would be enough to keep
the gain normally high. Habituation can take place only when no signals from
the tail occur for a long time. And of course habituation is not a phenomenon
in itself; it’s just a name for declining gain.
I have no guess about what the controlled variable is - it’s probably something
that has to do with the circumstances under which both mantle and tail are
being stimulated occasionally. Apparently the tail sensors have a rather high
threshold – unless there’s another neuron affecting signals coming from them.
I
would agree that the tail is probably being stimulated as aplysia is moving
about, but you may have overlooked something with respect to short-term
habituation. If you stop the train of repeated stimulation of the mantle
or siphon, the normal level of sensitivity slowly recovers. Based on what
little I’ve read about the cellular physiology involved, I imagine that
each touch of the mantle or siphon not only triggers mantle contraction (prior
to habituation), it produces some kind of transient deactivation of some
portion of the calcium gates in the terminal buttons. If you repeat the
stimulation soon enough, more deactivation occurs before the complete loss of deactivation
from the prior impulse. Consequently an increasing proportion of the
gates get deactivated, until there is no longer sufficient release of neurotransmitter
to fire the motor neuron or interneuron. A relatively long period of rest
allows the gates to recover, so habituation disappears.
Stimulation
arising from shocking the tail after habituation has been performed would
immediately reverse this cumulated deactivation of the Ca++ gates, producing immediate
dishabituation, as this form of recovery of the reflex is known.
BP earlier: I was looking on the web for more detailed
diagrams, but couldn’t find any – and none of them contains any information
about the transfer functions at each synapse. Aplysia contains about 20,000
neurons, so these little snippets don’t go very far toward giving us a picture
of the whole system
BP: The present article raises that number
to 100,000.
That
latter figure may be a mistake. The author of the paper you attached is a
biology professor; the paper is basically his lecture over the portion of the
textbook he is using that covers the neurology of learning. He’s not
involved in this research and is not an expert on aplysia. If there’s a
scientific paper claiming 100K neurons in aplysia, I haven’t seen that
source. But whatever the number, here we’re concerned with a portion that
numbers something like 24 sensory neurons and perhaps a similar number of
interneurons and motorneurons.
BA: Aplysia researchers
have been able to demonstrate what happens during habituation, both in terms of
changes in mantle contraction during repeated stimulation (typically once every
10 seconds in demonstrations of short-term habituation) and in terms of changes
in neurotransmitter release, post-synaptic potentials, and action potentials.
Concerns about whether there might be other inputs to this circuit that
influence the time-course of habituation led to in vitro experiments involving,
in isolation, only the neurons shown in the diagram I provided. Stimulation of
the sensory neurons produced the same pattern of changes found in the neurons
of the intact animal. The observed changes fully account for the changes in the
gill-withdrawal reflex during short-term habituation.
BP: The problem here is not seeing the
forest for the trees. In the Reiness article, the description drops to the
level of biochemistry and any Big Picture is rapidly lost. At the same time,
there’s that annoying habit that biologists have of casually introducing
purposive language at entirely too high a level of abstraction, but with quotes
so you know they don’t really, really mean “ignore because it was not
harmful.” Of course they don’t believe that Aplysia can perceive an
abstraction like harmfulness, or that it can direct attention toward or away
from some stimulus, or that it has “reasons” for what it does as
implied by the word “because.” But if they don’t believe that, why do
they talk that way? Especially about an in vitro preparation?
When
behaviorism was in its heyday, psychologists working with rats or other animals
used to put quotes around words denoting emotions (e.g., “fear”) to
alert the reader that this label is intended as shorthand for “behavior
that we humans might display if we were afraid, so something akin to this
experience might be present in the rat.” It was there to remind the
reader that we can’t get into a rat’s mind and experience what the
rat experiences, so we don’t really know whether that experience is
similar to our own. As time went on the quotes gradually disappeared, so now
cognitive psychologists don’t even acknowledge the possibility that the
rat may not have the same mental experiences that we humans do.
I
do sympathize with writers who are trying to communicate findings such as those
from aplysia research to students and other lay audiences. There is an
overpowering temptation to describe things to a lay audience in familiar terms.
Unfortunately, the audience probably will miss the fact that this language is
just a pedagogical device to facilitate communication, and will think of these
processes in exactly the terms used.
BA: Kandel and others
have shown that short-term habituation in this circuit depends exclusively on a
reduction in the amount of neurotransmitter being released with each impulse of
the sensory neuron. This reduction in turn results from decreased Ca++ intake
in the axon terminal during the action potential. Ca++ intake is
necessary for the transport of the vesicles containing the neurotransmitter
toward the synaptic gap. They’ve also identified the cellular changes
that reduce CA++ intake. All in all, they make a convincing case for the
mechanism of short-term habituation as found in the gill-withdrawal reflex.
Right, but somehow they missed the fact that the normal case has to be
maintenance of the amount of neurotransmitter release at the higher end of the
range – otherwise there would be no response to habituate.
See
above re: dishabituation.
BA: The shocks employed
produce localized effects in the tail. The reason shock is used is that it
meets the requirement of a stimulus that will produce sensitization. The input
must change suddenly and fairly strongly. Continuous tactile stimuli
wouldn’t work.
BA: Like something starting to take a bite
out of your tail. So the controlled variable is something like a rate of change
of stimulation of the tail and mantle. Shocks, however, do not produce
localized effects unless they’ve changed the laws of electricity. A shock to
your hand can stop your heart. Electricity goes everywhere it can go, which is
why I don’t like shocks as a physiological stimulus.
Actually,
a shock to the hand can’t stop your heart, unless the conduction pathway
to ground passes through the heart. Or so I understand. With respect to
aplysia, shock produces the same effect on the habituated gill-withdrawal
response as pinching the tail – a more natural stimulus, I’m sure
you’ll agree. Shock normally is used because it can be modulated
accurately and with precise timing.
BA: When it come to
neurologists speculating about what some collection of neurons in the brain may
be doing, I share your frustration. In this case, however, researchers have
taken the approach of identifying a relatively simple system that mediates a
specific reflex and has been shown to demonstrate both habituation and
sensitization, taken careful measurements of neural impulses, PSPs, changes taking
place in calcium gates and vesicle transport and so on, made observations on
reduced preparations in vitro and demonstrated that they reproduce those made
on the intact in vivo system, and so on. It seems to be precisely the sort of
work you’ve been looking for.
It is, but it’s masked by unnecessary complications and it stops short of being
useful – as usual. Either we get fanciful abstractions, or we’re drowned in
irrelevant (for purposes of understanding the circuit) details. And when you
finally start getting a handle on what’s happening, it turns out that they
missed the main fact – here, the fact that something must have been
maintaining the gain at a high level.
What
I’ve described here on CSGnet is only a small portion of the research
that has been conducted investigating nonassociative and associative learning
processes in aplysia. I’m starting to wade through some of it in preparation
for my Learning & Behavior text, but still have a long way to go before I
can say that I am familiar with and understand it. As the research progressed,
initial hypotheses and speculations about the circuitry gave way to other,
often more complex understandings. No doubt those ideas will have to be revised
further as new evidence comes in. However, the basic findings about what
changes at the synaptic and cellular level during the course of habituation and
sensitization seem to be holding up rather well. If anyone has access to
old issues of Scientific American, an early but quite nice description
of Kandel’s research on aplysia appeared in an article by Eric Kandel
entitled “Small Systems of Neurons” that appeared in September
1979, vol 241(3), 66-76.
More
recent research has demonstrated that what appears to be classical conditioning
can be obtained. This involves developing an association between a
previously neutral stimulus and the touch stimuli that elicit gill withdrawal.
The “association,” as expected, takes the form of changes in the
release of neurotransmitter at the synapse and changes in the number of
synapses between a given pair of neurons. Some of this research has been aimed
toward identifying whether the formation of this association follows the
Hebbian rule.
Bruce
A.