[From MK (2015.07.31.2340 CET)]
···
----
Subject: Re: Intrinsic reference conditions
[From Bill Powers (2007.12.24.0910 MST)]
Martin Taylor 2007.12.23.23.43
I guess I wasn't clear. An "error" in perceptual control
theory is the difference between a reference value and a
perceptual value in a control unit. The reason there is no
"intrinsic error" is that there is no reference value for an
intrinsic variable. If there is nothing for the value of a
variable to be compared against, the concept of "error" does
not apply.
[My comment continued]
On awakening this morning I got out B:CP and looked through it,
with some trepidation, for the discussion of homeostasis as it
relates to intrinsic reference signals and error signals.
Sure enough, it isn't there. Neither "homeostasis" nor "Cannon"
appears in the index nor, as far as I can find, in the text.
I was so focused on the connection between intrinsic error signals
and reorganization that I simply passed over the homeostatic
systems in which the reference signals and error signals appear.
I'm sure I must have written many times about homeostasis (I know
I reported to CSGnet upon discovering Mrosovsky's "Rheostasis"),
but I can't find anything about it in B:CP, even though I was
quite aware of that subject at the time of writing and considered
it to show a level of biochemical (and autonomic, as others have
reminded me) control systems. I can see that if another edition of
B:CP ever appears, it is going to require an added chapter on this
subject, or a large revision of the chapter on learning and
reorganization. I tell you, discovering a blind spot that large is
very painful.
One painful aspect of it is remembering how, when Gary Cziko wrote
about Bernard and Cannon in Without Miracles, I wondered why he
didn't credit me with applying control theory to homeostasis. The
reason is now quite clear: I didn't. I only thought I had done so.
So: my somewhat perfunctory mention of the possibility of a lack
of clear communication on my part turns out to be a very likely
explanation for why you. Martin, and probably many others don't
realize that the intrinsic control systems of which I spoke were
the same homeostatic systems that Bernard and then Cannon
recognized, and that led Arturo Rosenbleuth, a student of
Cannon's, to bring this subject to Norbert Wiener's attention,
thus giving rise to cybernetics. My only addition was to propose
that large enough error signals (how I wish I had termed them
homeostatic error signals) cause reorganization of the behavioral
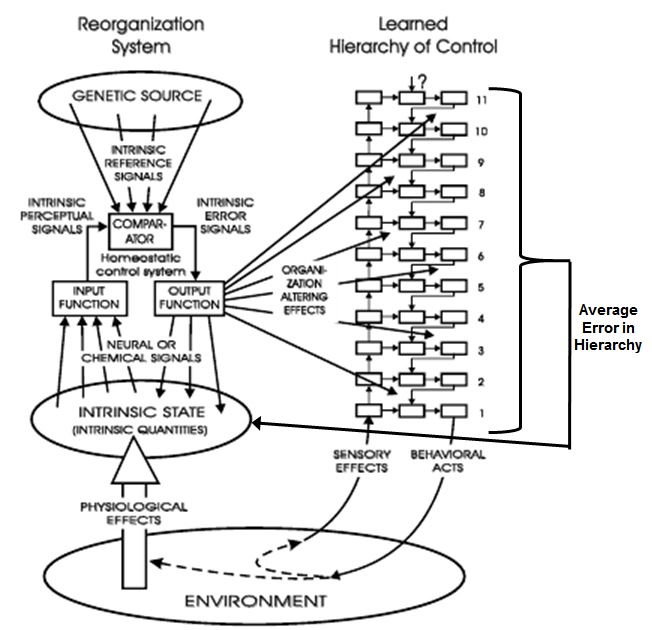
systems to begin. In my diagram of the relationship of the
reorganizing system to the behavioral hierarchy (Fig. 14.1) I show
ONLY the reorganizing effects of intrinsic error signals. The gap
left by omitting the local output functions that normally correct
intrinsic errors is now the most prominent feature of that diagram
in my mind. How could I not have seen what I was leaving out?
Dag Forssell, since it was you who drew the latest and clearest
version of Fig. 14.1, perhaps you could undertake to add those
missing output functions that convert intrinsic error signals into
physiological effects in that part of the diagram. But read on
first.
Writing this, I now realize that the "ignoration" of the
homoeostatic control systems was more than a simple omission. I
failed to see a principle that becomes obvious when the
homeostatic systems are added in all their glory as complete
control systems. When the physiological loops are added, we see
that reorganization is triggered by excessive and prolonged error
signals in somatic control systems -- just as it is triggered by
excessive neural error signals in the behavioral systems of the
brain. This quickly brings in another consideration that I have
looked at and mentioned, which is that "pain" in many cases (if
not all) is simply an ordinary perceptual signal that is excessive
in magnitude, meaning that it is causing very large error signals.
Any perception, when carried to an extreme magnitude, is painful -
- we try very hard to make it smaller. We can now say that any
error signal, whether in a biochemical, autonomic, or behavioral
control system, will, when large enough and protracted enough, be
experienced as pain and will cause reorganization to begin.
This tells us that the reorganizing system must be a distributed
system that brings reorganization to all levels of control systems
from bottom to top. At the level of DNA, it exists in the form of
repair enzymes. The immune system is a higher-order version of
repair enzymes. Reorganization exists at every level and acts
locally to that level. So we arrive at the question, "what about
amoebae?" And the answer, too.
Reorganization is simply an aspect of any level of biological
control systems.
And that brings up a realization delayed by some 35 years because
of that blind spot: every level of organization has ITS OWN
reorganizing system that senses excessive error and applies its
reorganizing actions to that level. So the diagram of Fig. 14.1 is
probably wrong. It is not error at the physiological level, but
only error at the behavioral level, that leads to reorganization
at the behavioral (neural, brain) level. Reorganization does
result from excessive error at the homeostatic level, but its
effects happen at that level. If we reorganize our behavior
because of physiological problems, we do so only because those
physiological problems are not corrected by reorganization at the
physiological level, and lead to excessive errors in the
behavioral systems. It is the latter kind of error that leads to
reorganization at the behavioral level. So now we see that every
new level has to deal with whatever errors the levels below it
can't handle, with reorganization happening just as control of any
kind happens: locally.
I don't know how well this revision will survive aging, but it's
pretty clear that it wouldn't have occurred to me if you, Martin,
hadn't made the inflammatory proposal that there are no intrinsic
reference signals.
Best,
Bill P.
----
M