···
On Thu, Sep 3, 2015 at 12:20 AM, PHILIP JERAIR YERANOSIAN pyeranos@ucla.edu wrote:
[philip 9.2.15]
music starts playing
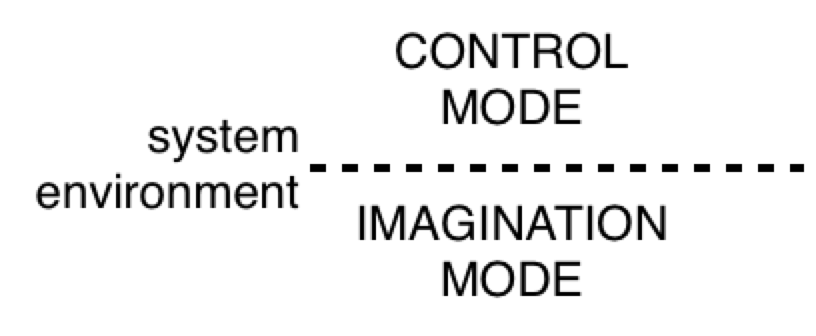
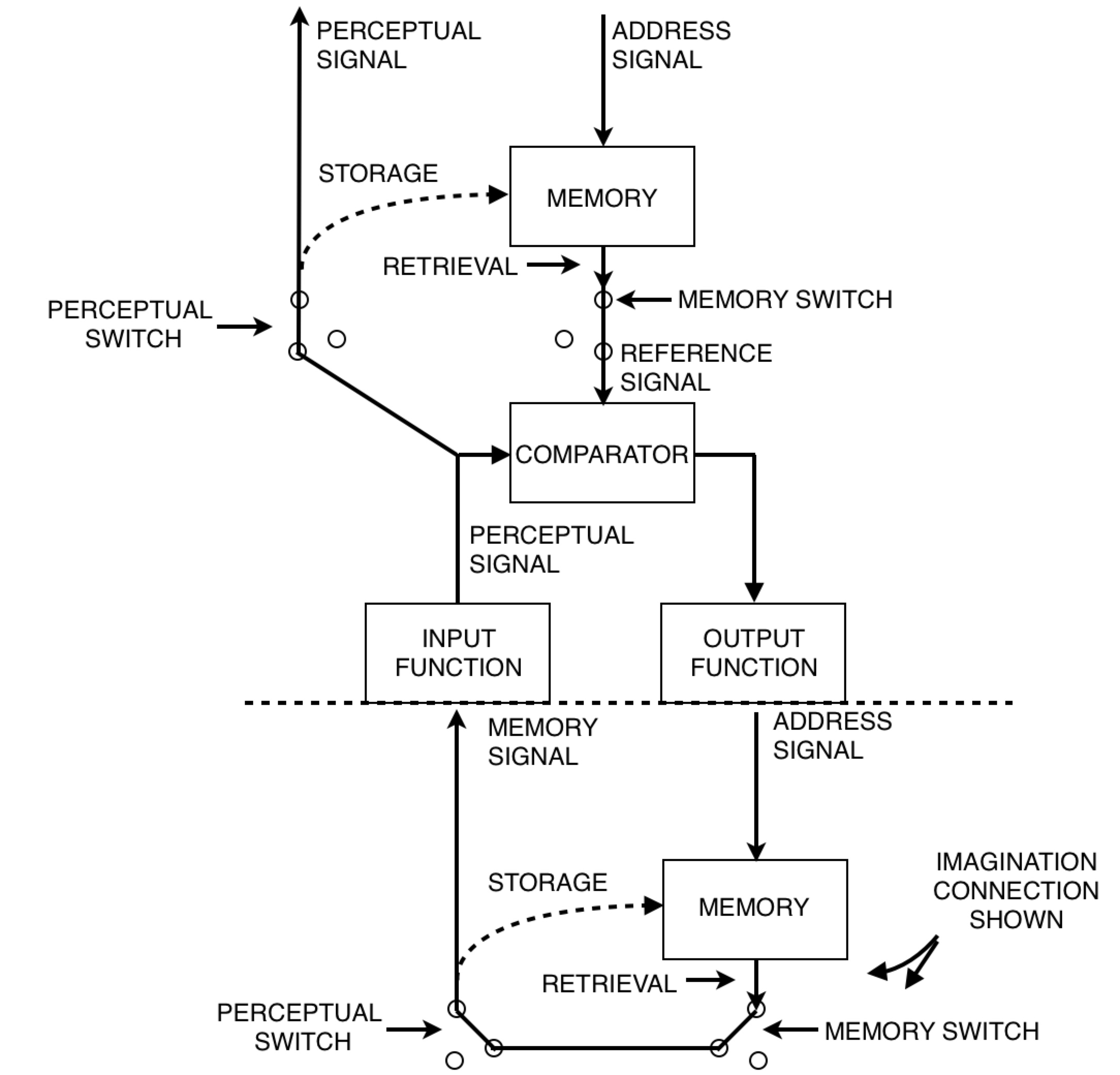
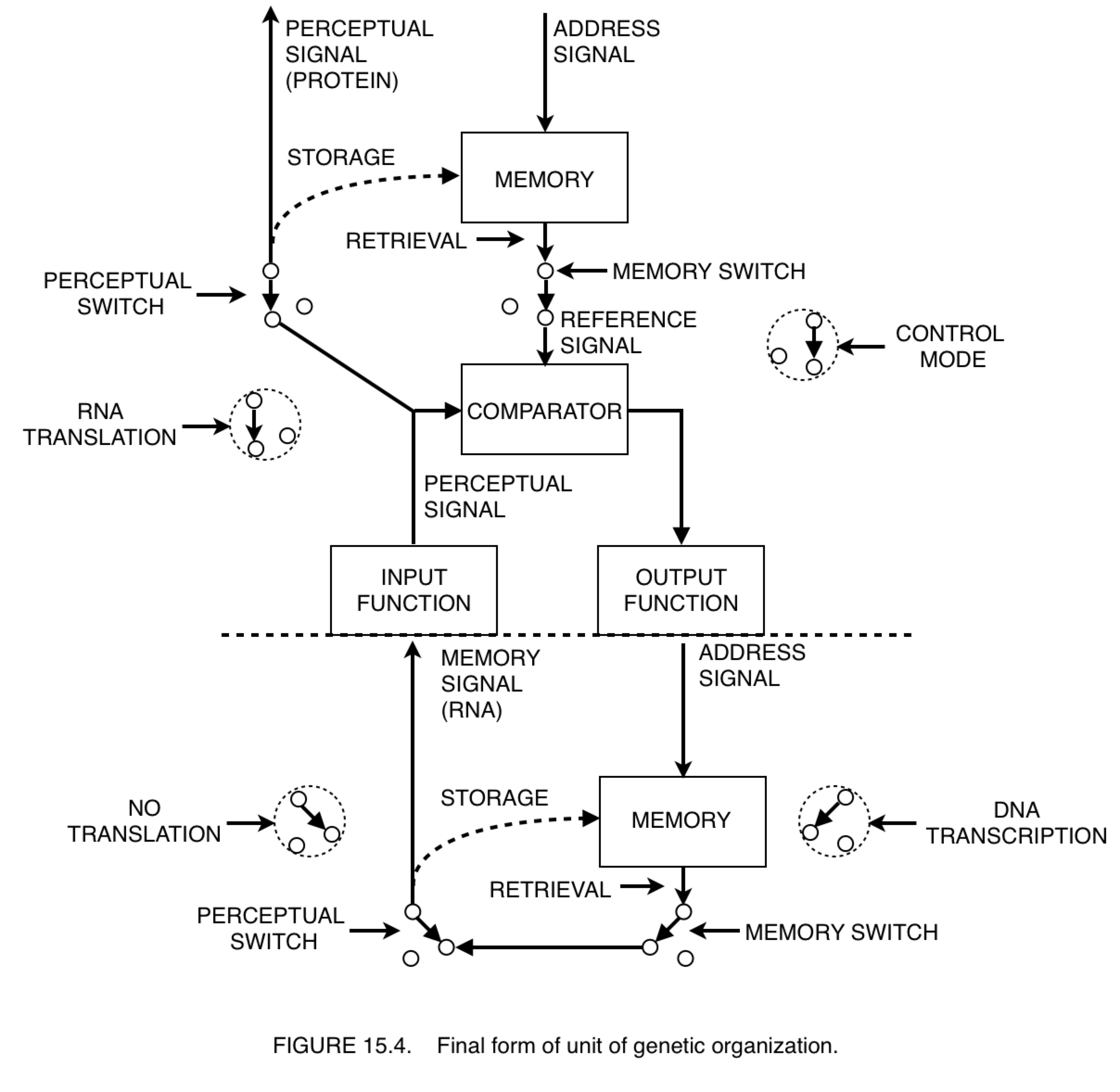
And we're back from commerical break! Let's start with a talk about B:CP. I've been hearing a lot lately about the difference between control of perception and imagination. This is a great topic. Take a look at Figure 15.3 on page 221 (powers, 1973). Observe carefully where it says "imagination connection shown".
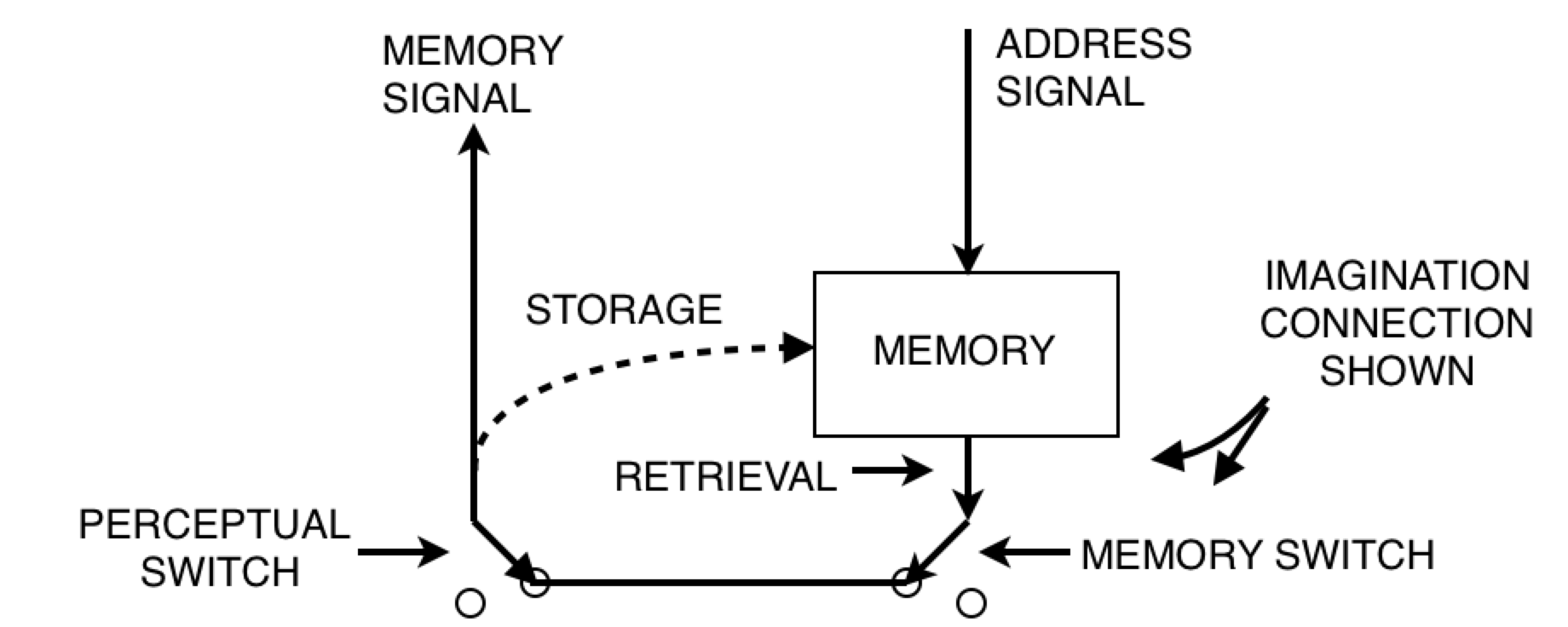
On page 210, Powers says, “There are no lower-order sources of memory for first-order systems to perceive; hence the lowest level of memory content should be sensations.” Well, I don’t think there’s proper grounds not to allow this memory signal above to be perceived by a first order system. So let’s allow it. After all, we want to allow the model to build computers, and the diagram above is what a computer looks like! So where does this leave us?
In a nutshell, “control of imagination” means this:
We have a system in control mode above a system in imagination mode, and the controlled quantity is a memory signal. I suppose one might argue that the only thing we can do in imagination mode is to remember what we imagined. But can anyone tell me how to quantify memory? The punchline is yet to come, along with Figure 15.4. Stay classy, my friends.
cut to commercial
On Wed, Sep 2, 2015 at 7:45 PM, PHILIP JERAIR YERANOSIAN pyeranos@ucla.edu wrote:
[philip 9.2.15]
Pardon me, the last message was shot off prematurely.
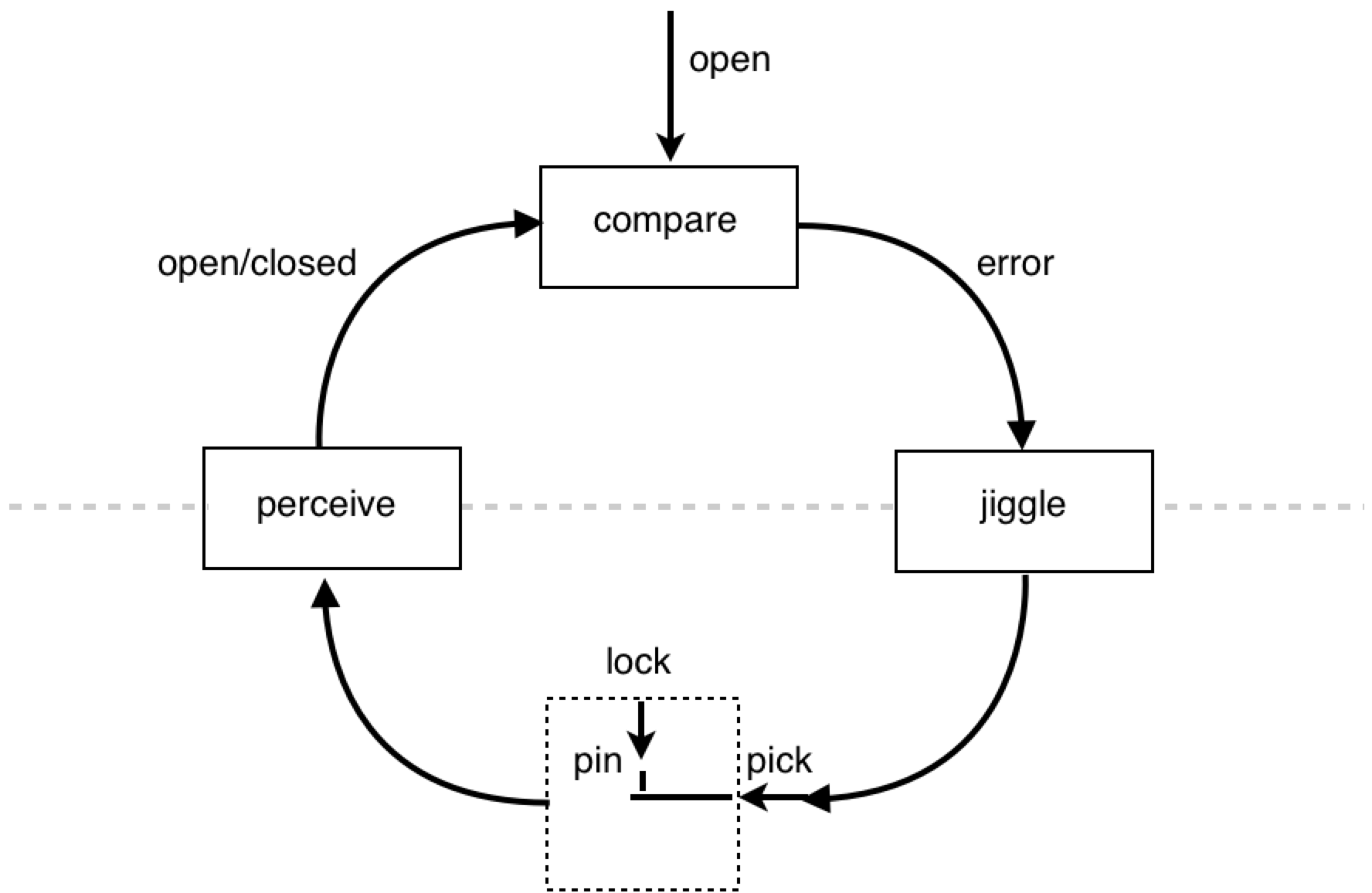
Anyway, today we’re going to talk about lock picking.
In case you haven’t read the book on lock picking, it’s OK, I will explain the concept behind the ordinary pin-tumbler. Peep into the key hole of a lock and you’ll see darkness. But if you stare long enough, you might see the first of an array of pins. These pins are there to prevent someone from opening the door without a key.
front view (ordinarily visible)
side view (ordinarily hidden)
Keys usually have a numeric code printed on them (for key makers’ reference) specifying the shape of the key. The key shown above has the code 3 1 2, because the successive ridges on the key are of respective heights 3, 1, and 2. These numbers correspond to the height which the pins in the lock need to be raised. We don’t need to go into the details, but you can open a lock without knowing the key code. You simply use a lock picking device as an analogue for a key and you “wiggle” or “jiggle” the pick (“jiggle” is the professional lingo). All you’re doing is varying the position of the pins in an unsystematic manner many times and relying on chance to align the pins. Note, there is also a more systematic method of lock picking, but it involves the same concept of varying the positions of the pins until they are at reference levels.
Now, the main reason I wanted to talk about the pin tumbler is because I think it should be the quintessential example of a perceptual control system interacting with its environment. It’s a very simple example, and it’s interesting as a PCT exercise because it involves interacting with the environment (the lock) through a device (the lock pick). On a side note, I used to wonder whether the perceptual control system organization is a lock. Today, however, I figured it would be a better idea to put it “on the other side of the dotted line” - i.e. in the environment.
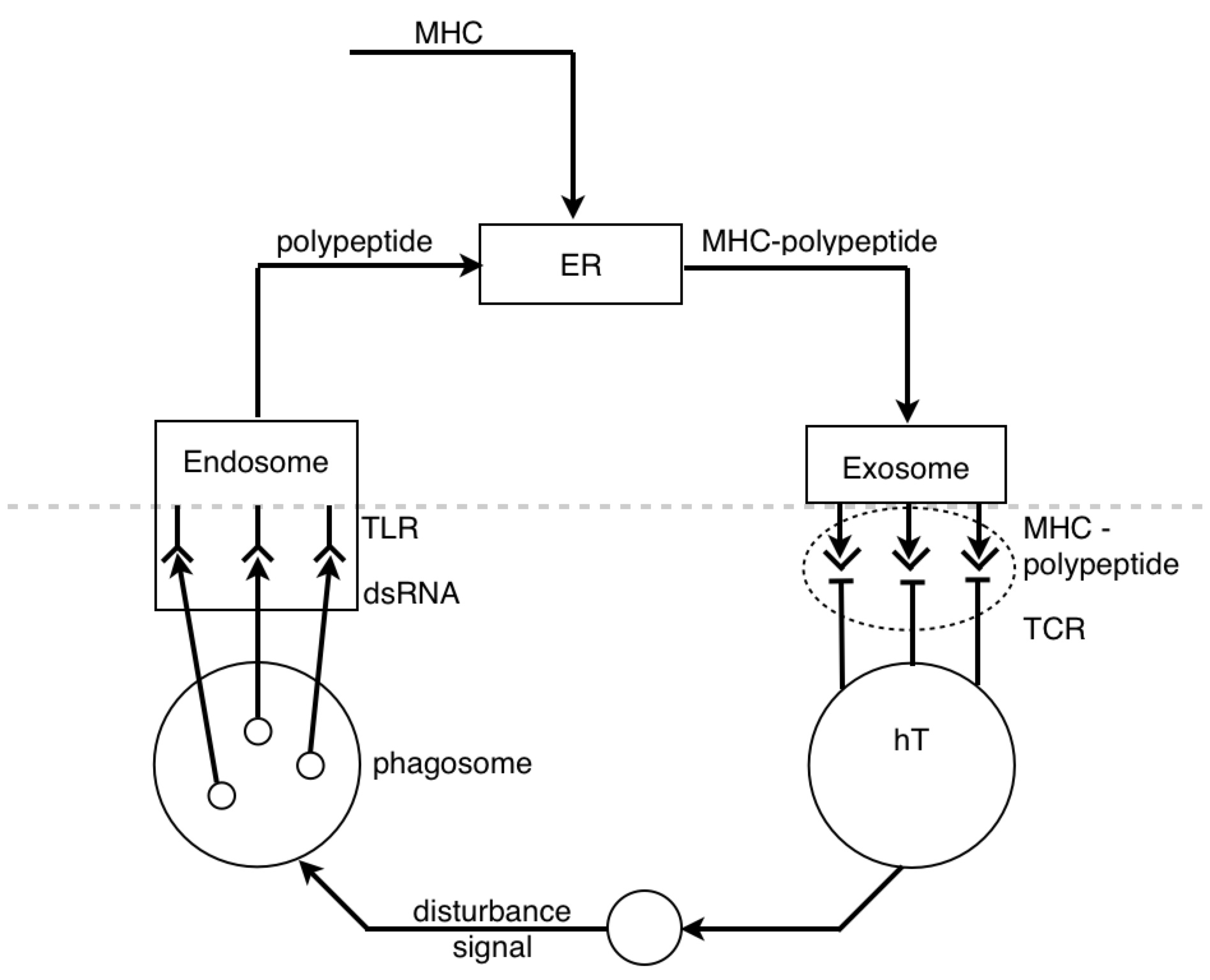
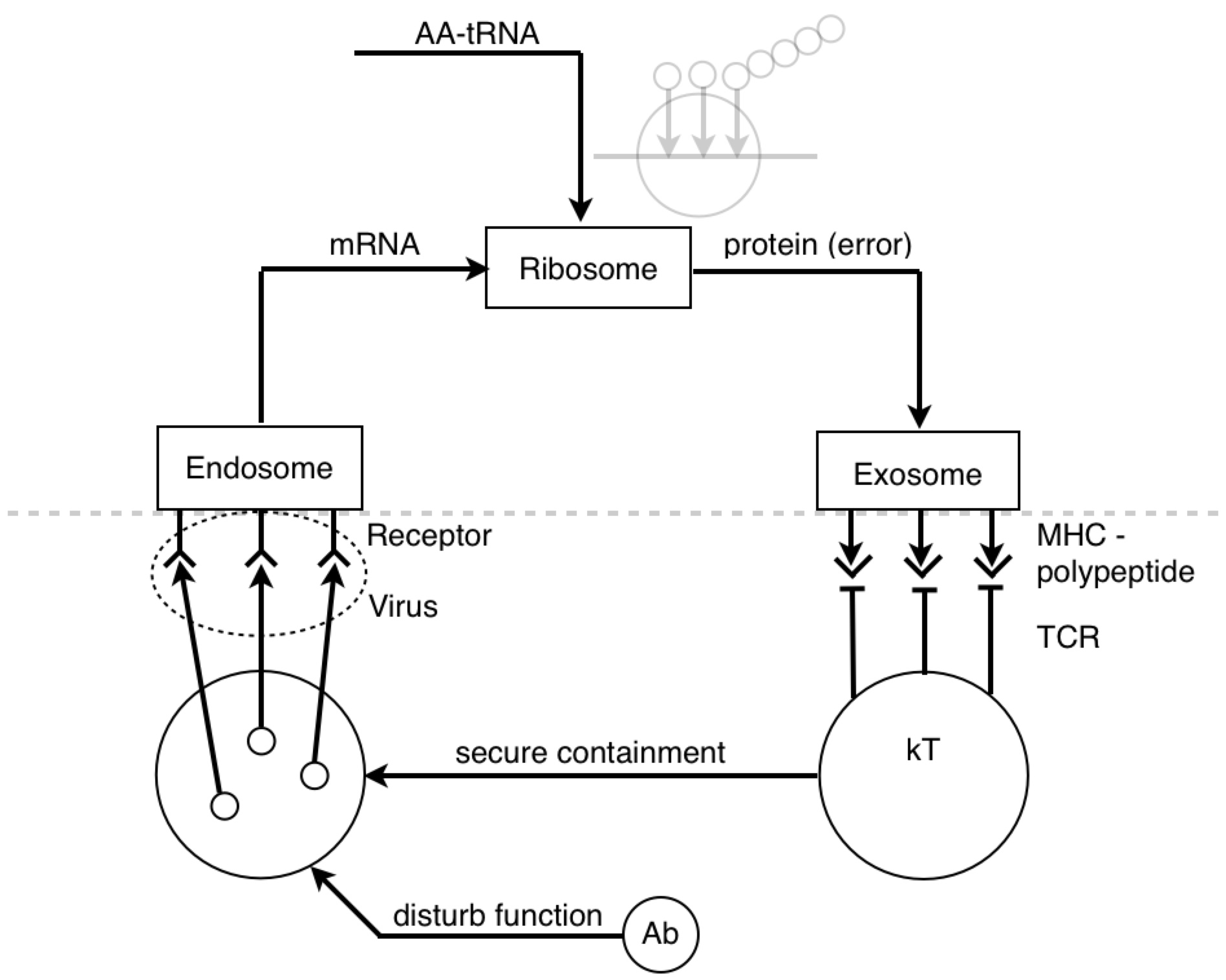
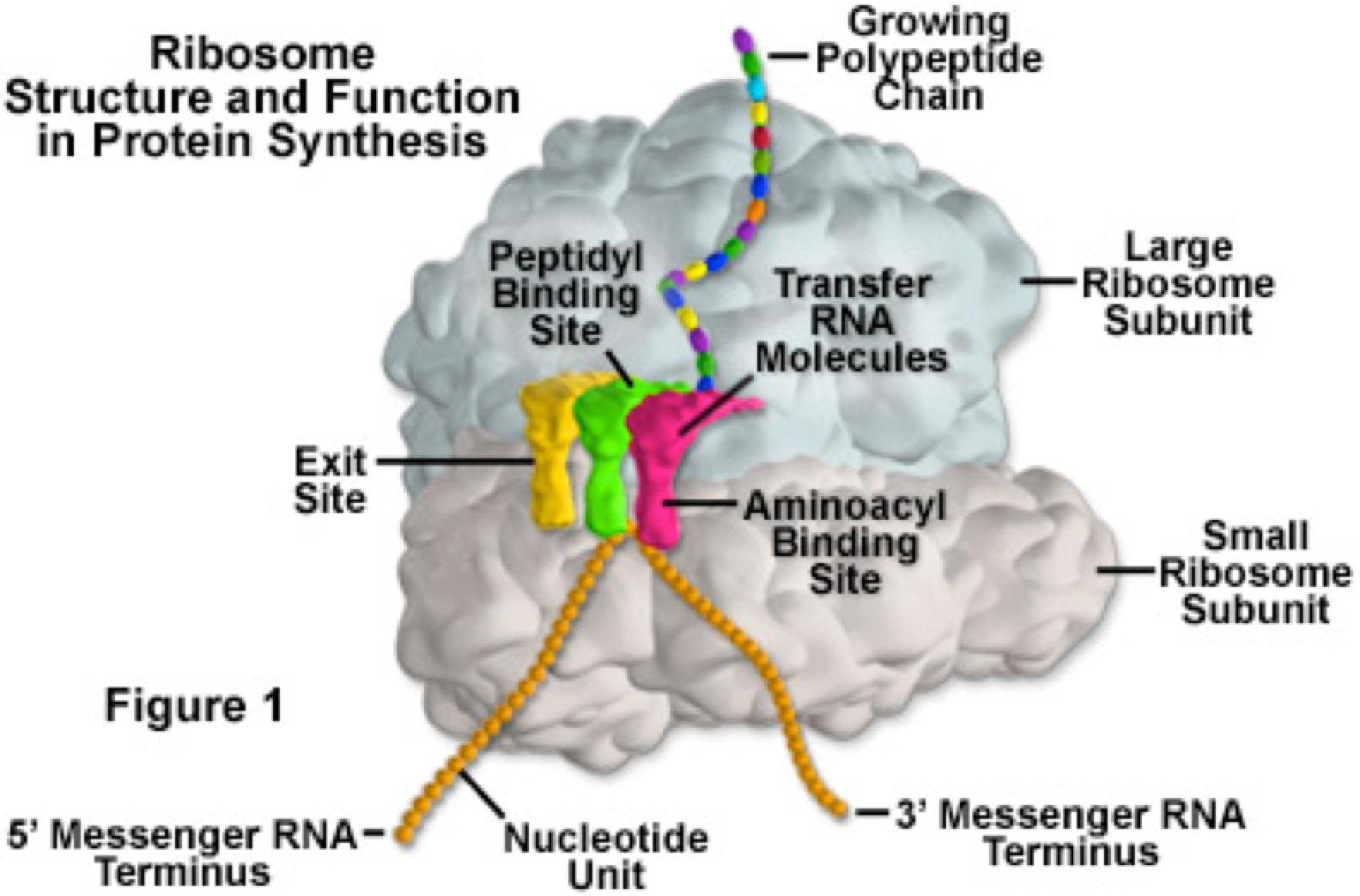
And the other reason I wanted to talk about the pin tumbler is because, when I look at a picture of the ribosome I see the pin tumbler.
I know it’s being used as a “translating” device.
But where exactly does it fit into a PCT diagram? I’ve never seen a lock in a PCT diagram before…until today.
music starts playing
Up next, we talk about genetics and epigenetics. Also, find out how you can save 15% or more on your behavioral psychology insurance.
On Wed, Sep 2, 2015 at 4:32 PM, PHILIP JERAIR YERANOSIAN pyeranos@ucla.edu wrote:
[philip 9.2.15]
Today we’re going to talk about lock picking.
On Wed, Sep 2, 2015 at 1:18 AM, PHILIP JERAIR YERANOSIAN pyeranos@ucla.edu wrote:
[philip 9.1.2015]
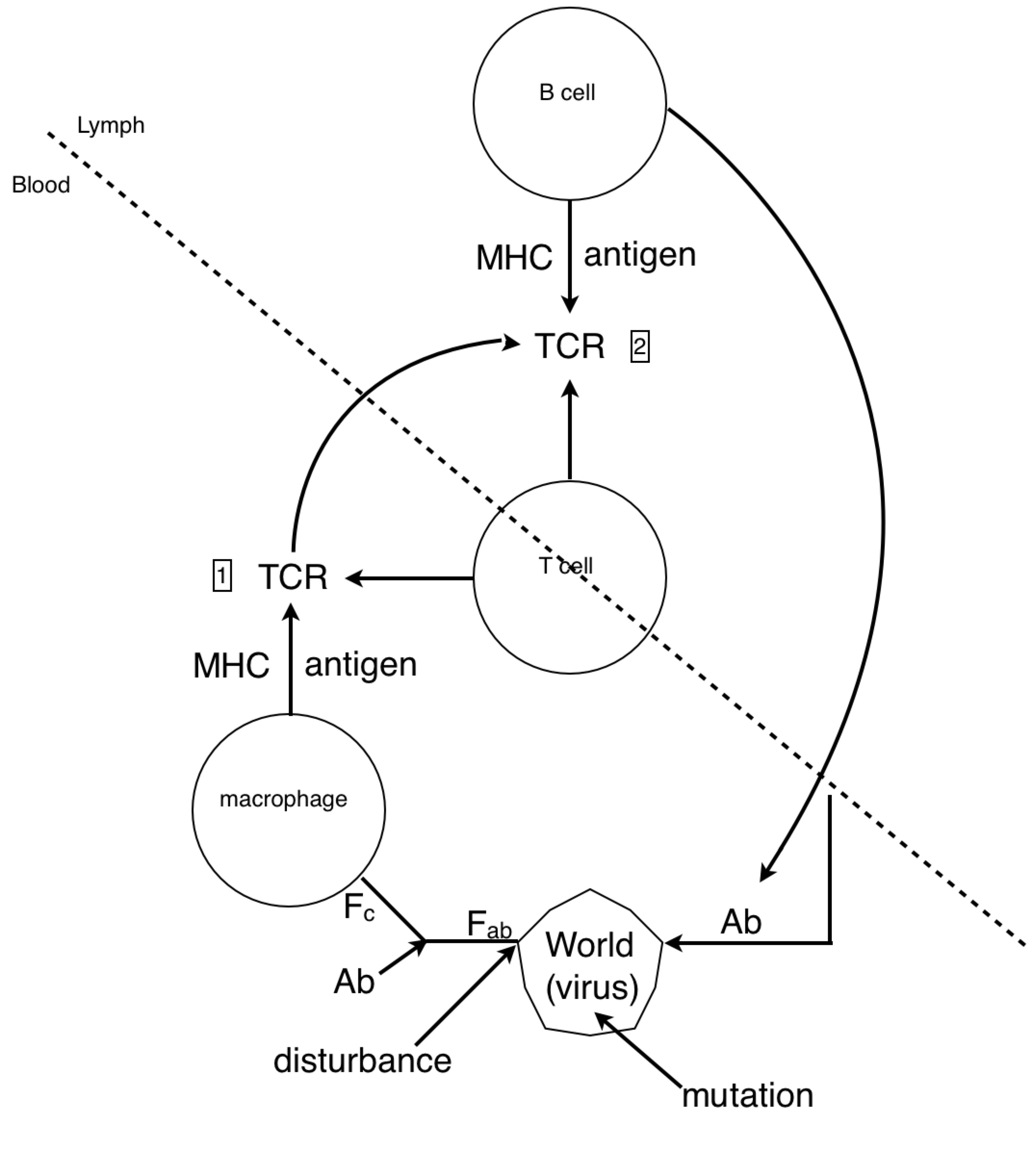
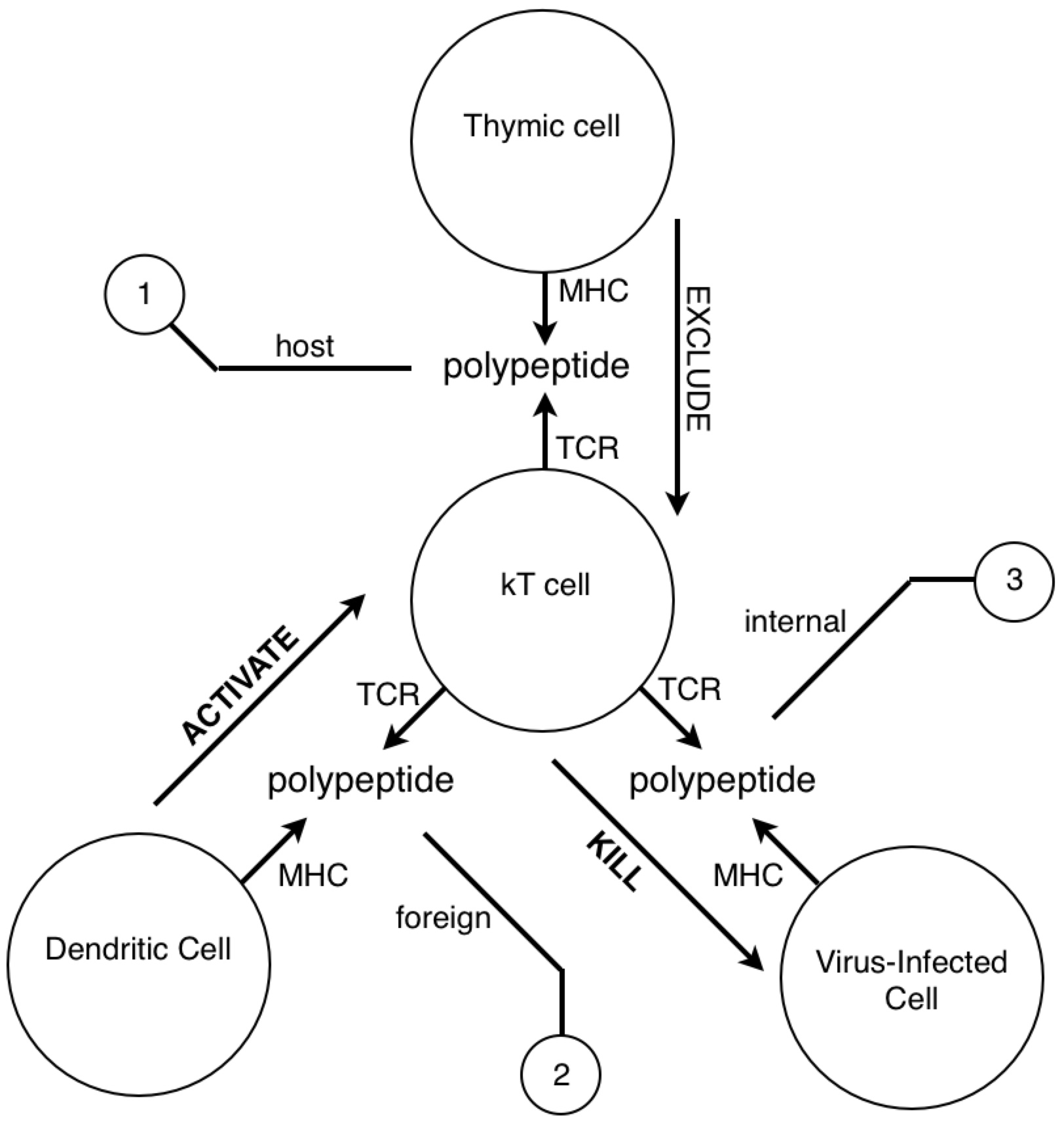
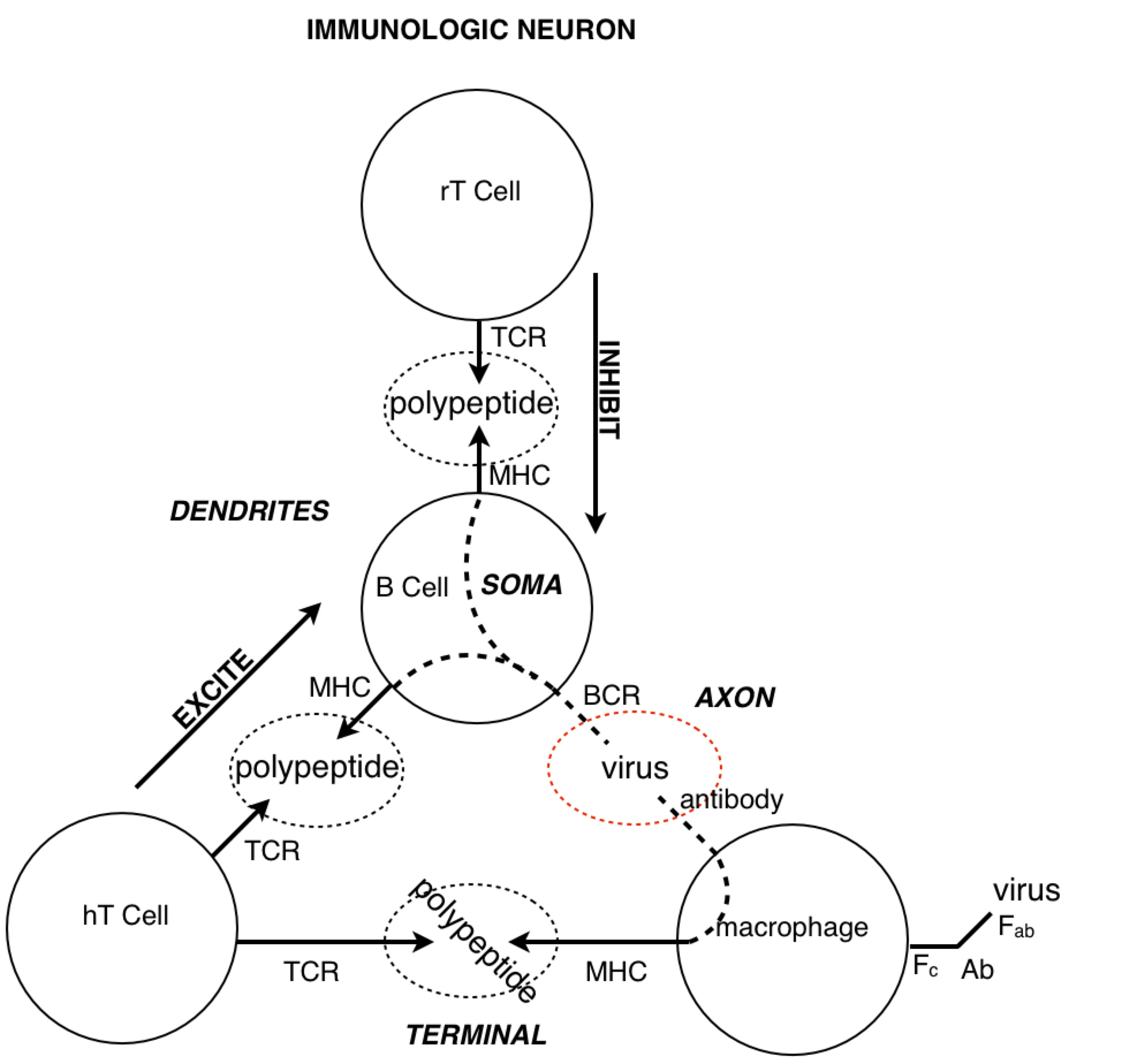
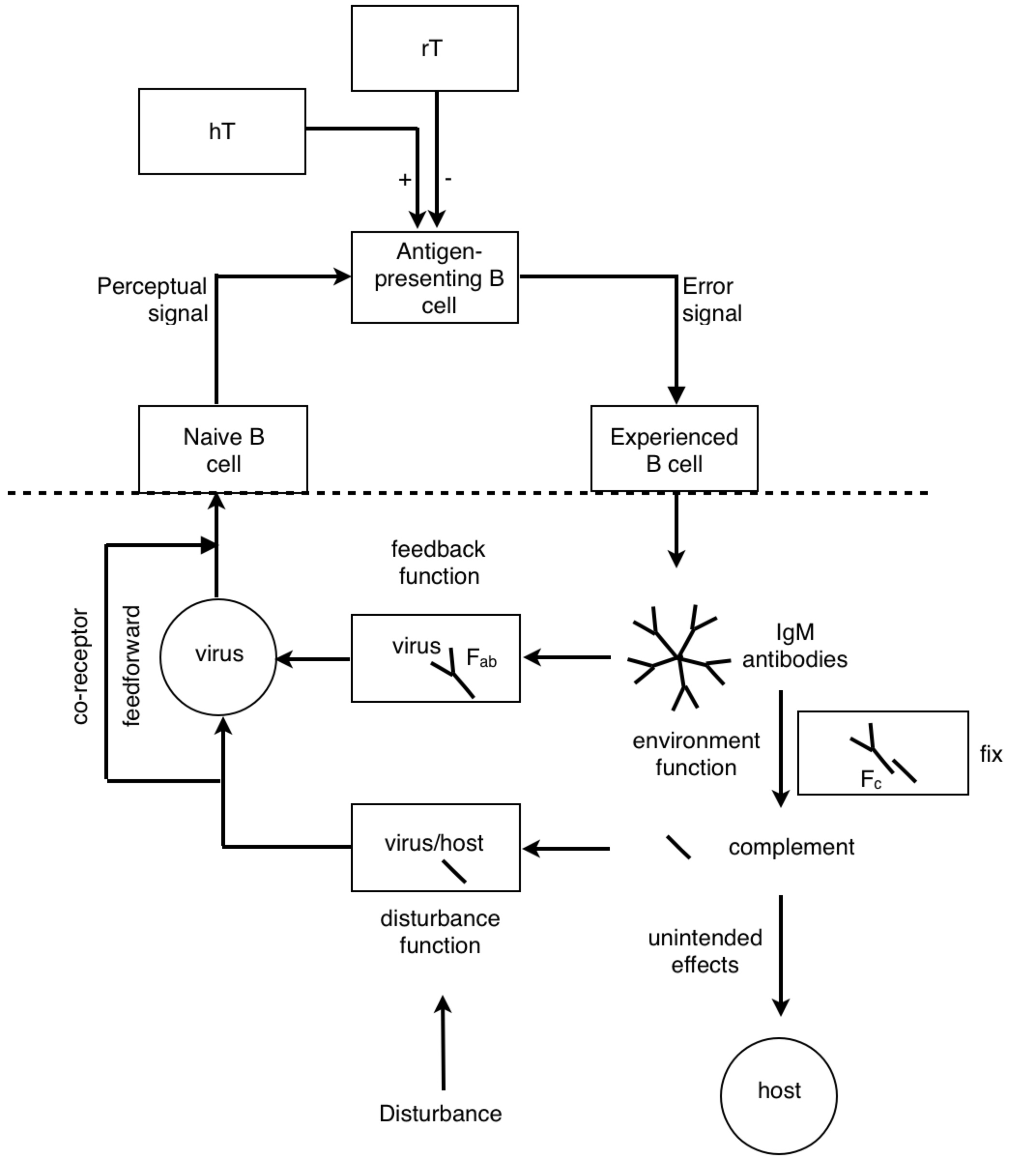
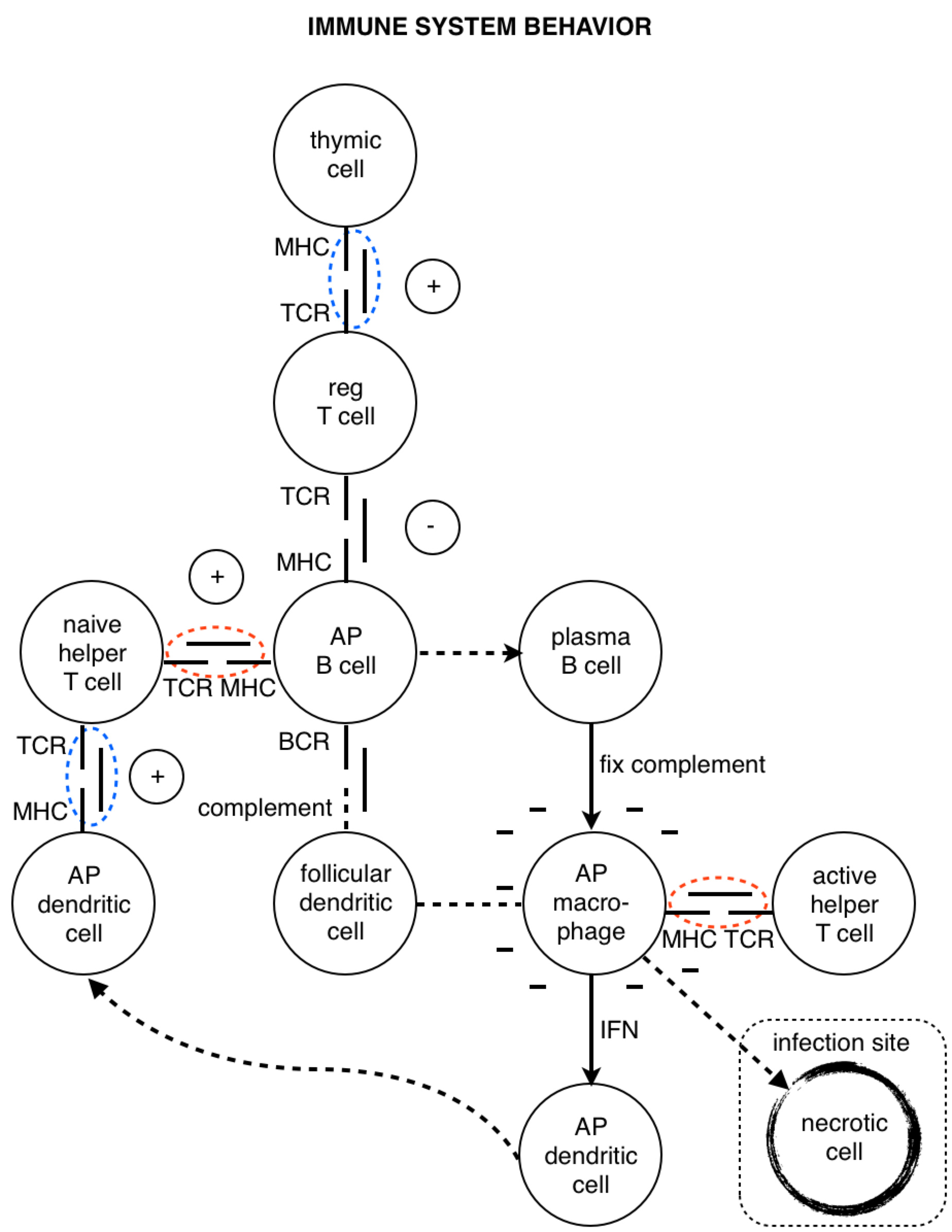
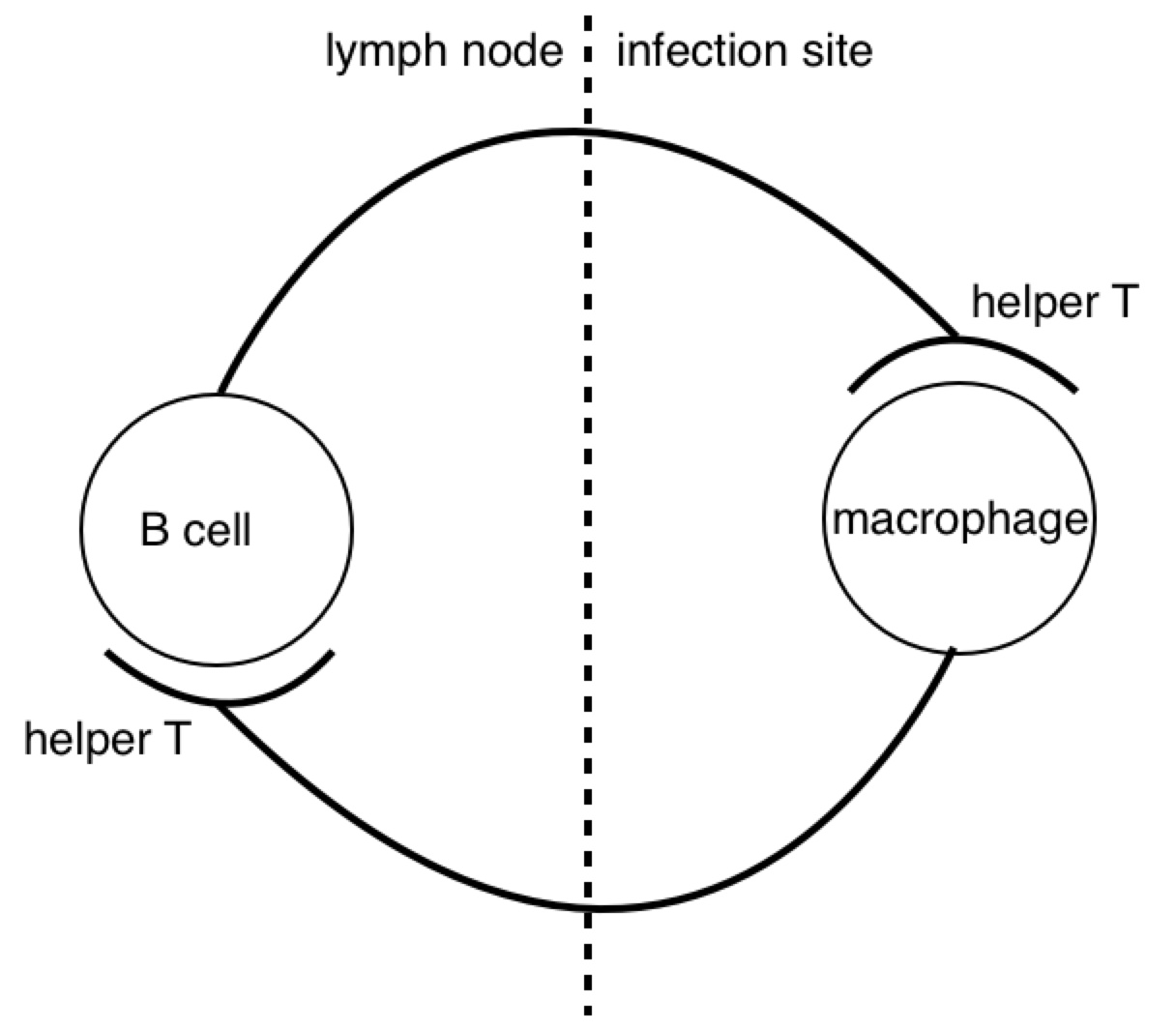
Look at how all the immune system players work together. We see the star player is the macrophage. At the start of the game it travels to an infection site where it’s activated by necrotic factors being released by dead cells. When it’s in place, activated macrophages generously express complement proteins (which I’ve depicted as dashes sprinkled around the macrophage).
The complement proteins are actually doing most of the killing, tearing holes in bacterial membranes left and right - killing them on the spot. Then, all these complement-opsonized shreds of bacteria either find themselves eaten by macrophages, or picked up by follicular dendritic cells in the lymph nodes.
If the macrophage eats the bacteria, it reorganizes the bacterial proteins and then represents them on its surface for helper T cells. But how would a helper T cell know what to look for? Aha! In the heat of battle, the macrophage dispatches nearby dendritic cells to transport a cargo of the bacterial protein to the lymph node, where the same reorganized proteins are represented to naive helper T cells.
If a naive T cell encounters its cognate antigen, it proliferates. Some of these T cells stay in the lymph and some travel to the blood, where they actively seek out infection sites. The traveling T cells are searching for macrophages which are presenting their cognate antigen; and once they find them, wherever they are, they keep them there by pumping out a diverse profile of signalling molecules which maintain the macrophages in hyperactivated states.
When an active T cell encounters its cognate antigen at an infection site, it proliferates again. Some of the cells stay at the battle scene, while others travel back to the lymph nodes to activate antigen-presenting B cells. Meanwhile, the complement system has been dumping tons of opsonized bacterial protein into the lymph nodes. This provides ample opportunity for any B cells to find their cognate antigen in time for the return of the battling T cells.
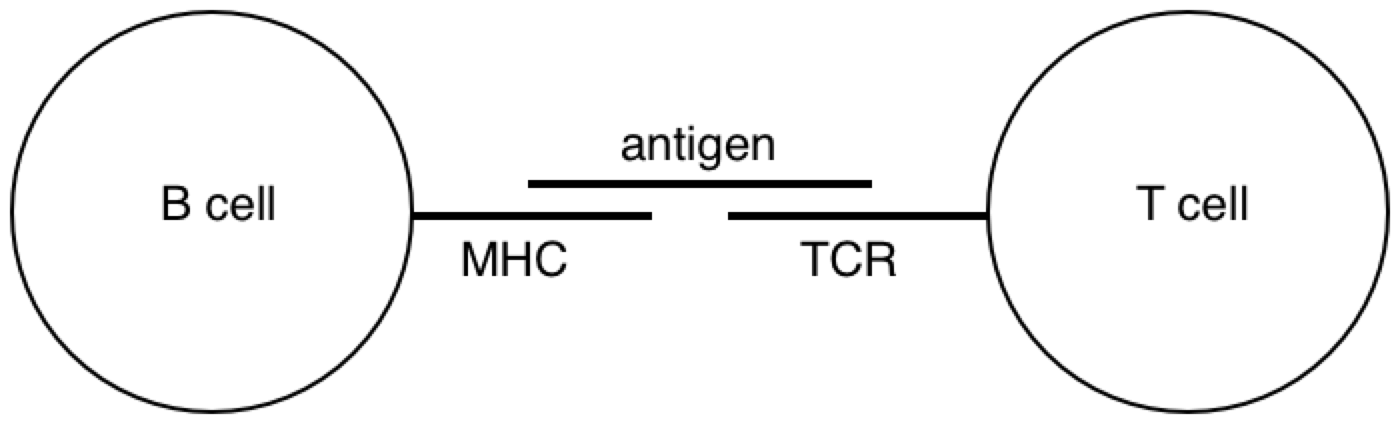
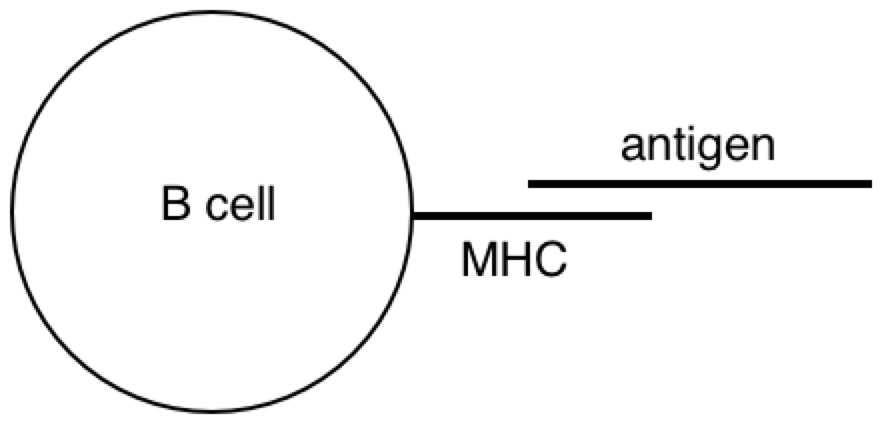
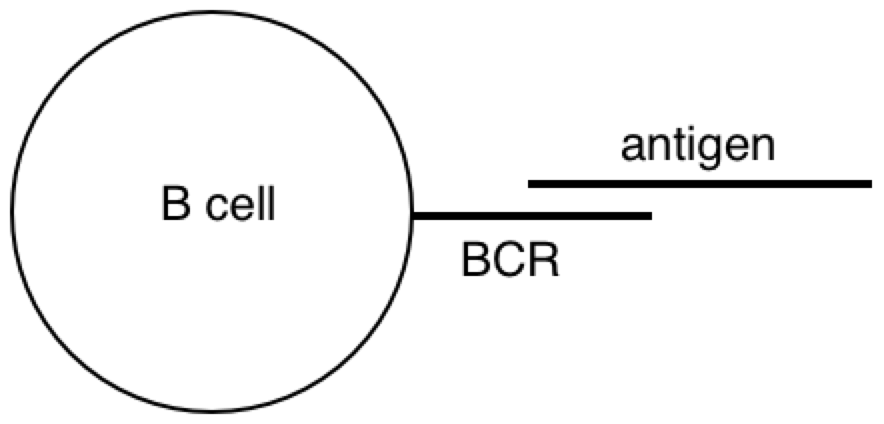
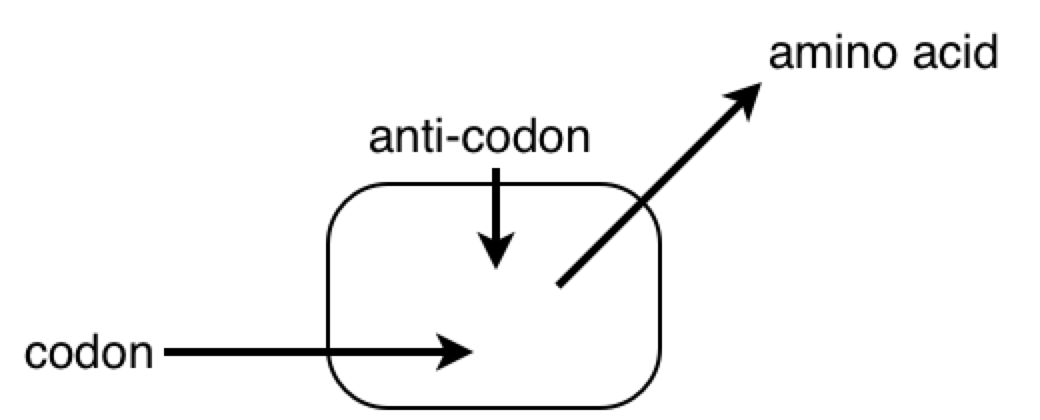
All this time, the B cells have been fishing for antigen swimming among the follicular dendritic cells. And those B cells with the highest affinity for the bacterial protein, compared with their neighbors, are going to have picked up the most antigen for presentation to T cells. Now the B cells pick up antigen with their BCRs, which are essentially like hooks (they bind to 2º structures, like loops). In contrast, TCRs are essentially like keys (they bind to 1º structures, like a sequence). So as it turns out, the T cell is basically looking to the B cell in the lymph node for the same amino acid sequence it saw from the macrophage at the battle scene (refer to the parts of the above diagram circled in red).
The antibody doesn’t make it to the battle scene until all this is over. But once those activated B cells move out of the lymph and start pumping out antibodies into the battle scenes, it’s pretty much all over (for the bacteria).
Now it might not be obvious, but the immune response is made up of a bunch of motifs which look like this:
A closed loop of neurons. We notice a synapse between helper T cells and macrophages and between helper T cells and B cells. Recall that both B cells and macrophages are antigen presenting cells for T cells. Now, if we pay close attention to the proliferation and migration patterns of T cells, we’ll see that helper T cells proliferate before encountering macrophages, and once more before encountering B cells. So during an active infection, when there is plenty of antigen going around, we see many of these closed pathways forming. But when the infection subsides, and the antigen levels run low, we see many of these synapses go away. This process seems analogous to potentiation and depotentiation in neurons.
That’s all for tonight’s lesson. Tune in next time.
On Tue, Sep 1, 2015 at 2:47 PM, PHILIP JERAIR YERANOSIAN pyeranos@ucla.edu wrote:
[philip 9.1.2015]
WM: Agreed! Philip, Do you have a publication outlet in mind when you are happy with it?
PY: LCS IV
RM: This is a little more comprehensible to me, given my very meager understanding of biochemistry. I think the system above the dashed line is a lymphocyte; the stuff below the line is the environment
of the lymphocyte (the blood?).
PY: The lymph 
RM: The lymphocyte is controlling something
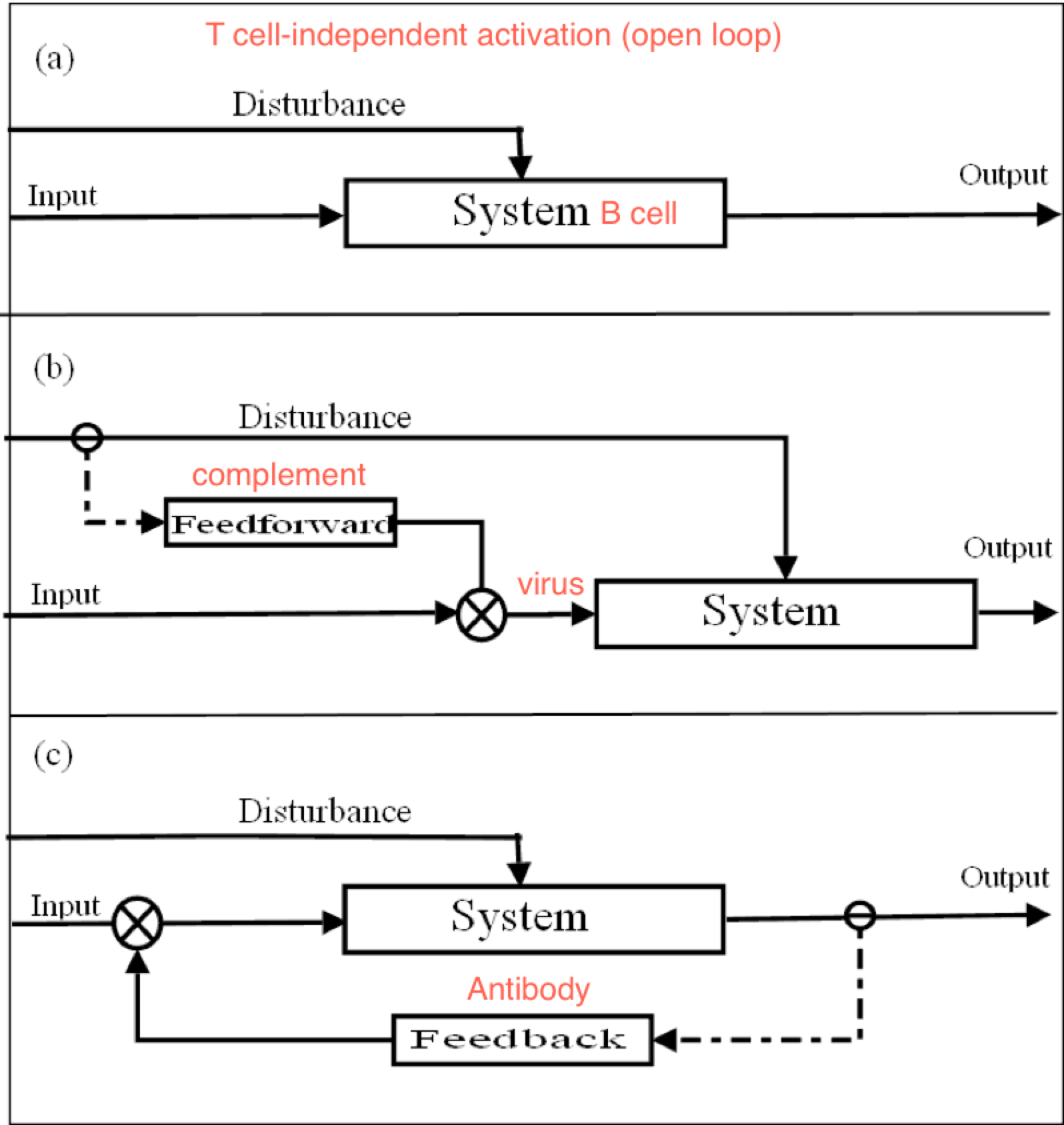
about the virus. Since, as I understand it, lymphocytes eliminate viruses, or try to, the lymphocyte is controlling for getting whatever it perceives about the virus to zero. And it does it by producing antibodies. So what I see here is an attempt to model the control of viruses by lymphocytes and basing the structure of the model on what is known about the interactions between biochemical variables in the lymphocyte and its environment. Is that right?
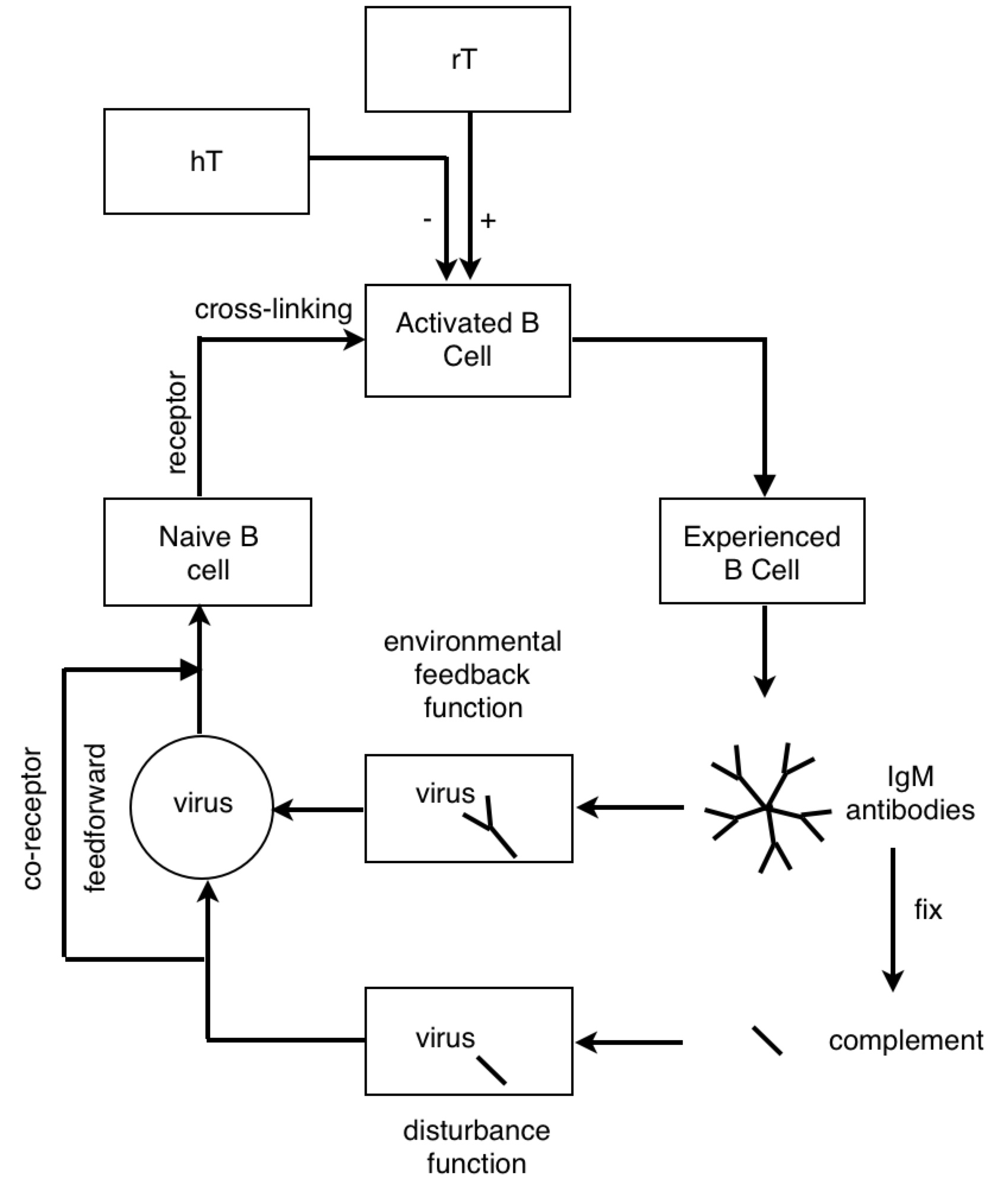
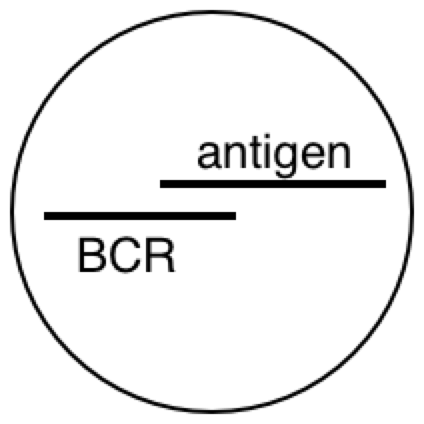
PY: The B lymphocyte is perceiving something about the virus with its BCR, and the T lymphocyte is perceiving something about the virus with its TCR. At some point, the antigen goes through the B cell and pops back out before hitting the T cell.
PY: Now I will argue that the antigen-presenting B cell is a reorganizing system.
BP: [reorganization]: The process of changing the forms of functions in the
hierarchy of control systems. see [intrinsic error]
[intrinsic error]: A discrepancy between any intrinsic quantity and its
intrinsic reference level. The driving force for reorganization.
PY: Recall, reorganization is a process caused by a discrepany between an intrinsic [physical] quantity and its intrinsic reference level.
BP: [physical quantity, phenomenon]: A perception identified as part of a
physical model of [external] reality.
[external reality]: A directly-perceived set of hypotheses, beliefs,
deductions, and organized models purporting to explain directly
[perceived] reality in terms of underlying phenomena and laws.
[perceived reality]: …the subjective impression of a three-dimensional
outside universe.
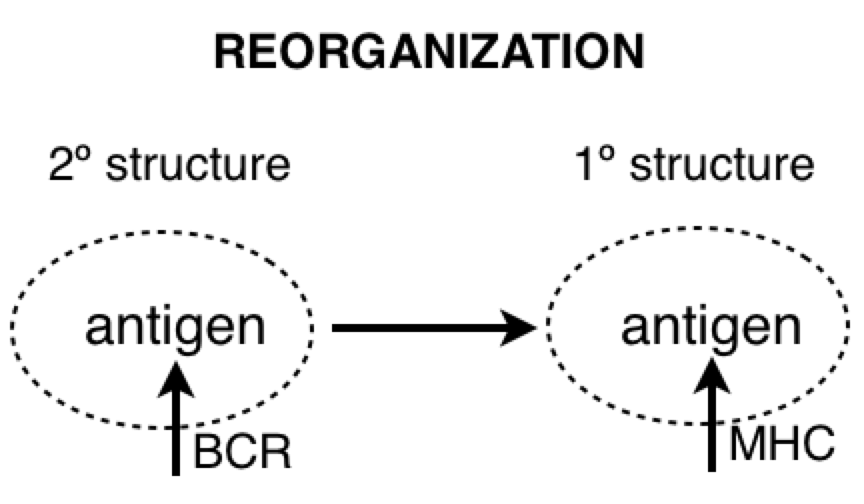
PY: Ok…The control hierarchy for microbiological physics goes 1º (primary) structure controls 2º (secondary) structure, etc. And data and logic suggest that primary structures are more numerous than secondary structures - with many different 1º structures (sequence permutations of an array of amino acids) having equivalent secondary structures (3-D shape of polar/non-polar amino acid interactions). So if we see a transition from detecting a 2º structure to a 1º structure, then we might suppose this is an information-generating process. And we might suppose that the process of changing the form of functions or structures of the hierarchy is reorganization.
RM: The lymphocyte is controlling for getting whatever it perceives about the virus to zero. And it does it by producing antibodies.
PY:
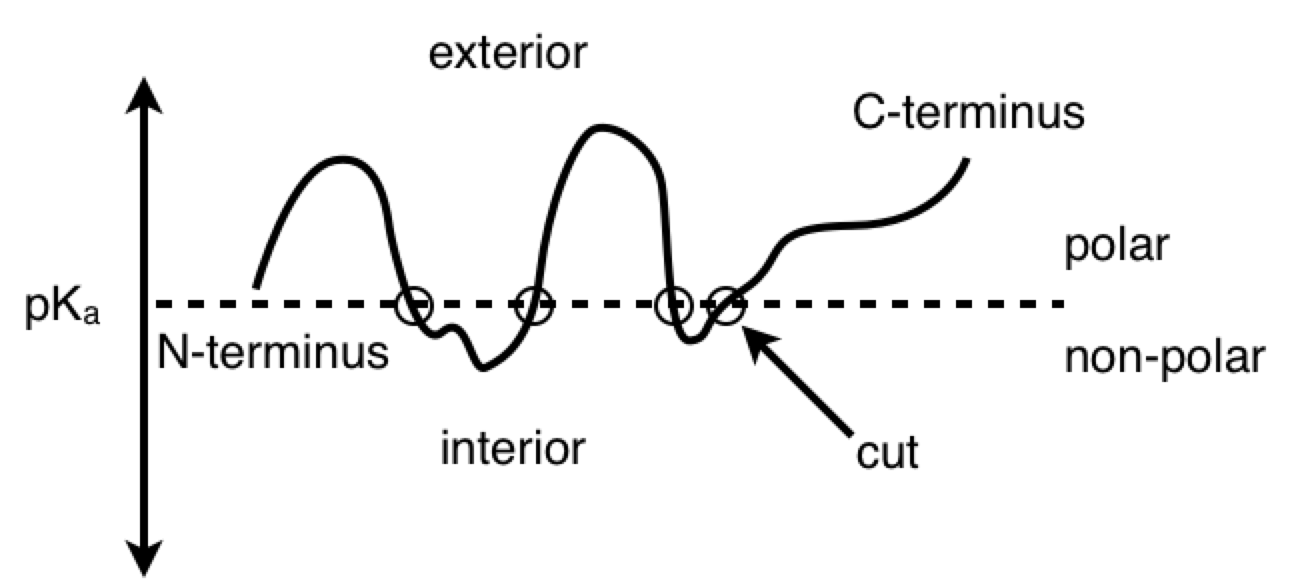
PY: Here is a diagram of the virus protein. It has polar and non-polar amino acids, which are going to associate with polar and non-polar regions respectively. The BCR is obviously going to interact with some hydrophilic binding motif on the exterior of the protein (instead of some hydrophobic region tucked away in the protein interior). When the virus protein enters the antigen-presenting B cell, the BCR passes the protein along to the proteasomes. And as the proteasomes are scanning the surface of the protein, they preferentially cut the C-terminal at non-polar, hydrophobic amino acids. This allows the BCR binding sites to remain generally unaltered. So, in a strange sense, the lymphocyte is indeed controlling for getting whatever it perceives about the virus (some hydrophilic binding domain) to zero (no perception). And this allows, some 2º structure detected by the BCR to be reorganized into its 1º structure for detection by the TCR.
RM: So what I see here is an attempt to model the control of viruses by lymphocytes and basing the structure of the model on what is known about the interactions between biochemical variables in the lymphocyte and its environment. Is that right?
PY: Yes, that is right. Let’s recap: the BCR’s antigen binding region’s shape is a subjective impression of a 3-D outside universe - a perceived reality (2º structures). This reality is explained in terms of directly perceived underlying phenomena and physical quantities (1º structures). This is done by reorganizing or changing the form of the protein structure inside the B lymphocyte, and then detecting the reorganized structure.
RM: Anyway, the diagrams are really nice, even if I don’t understand them. And you are to be commended for all the hard work you’ve put into this.
PY: I know I said I wasn’t asking for any compliments… but thank you
On Tue, Sep 1, 2015 at 9:02 AM, Warren Mansell csgnet@lists.illinois.edu wrote:
Agreed! Philip, Do you have a publication outlet in mind when you are happy with it?
Warren
On Tuesday, September 1, 2015, Richard Marken csgnet@lists.illinois.edu wrote:
[From Rick Marken (2015.09.01.0755)]
–
Dr Warren Mansell
Reader in Clinical Psychology
School of Psychological Sciences
2nd Floor Zochonis Building
University of Manchester
Oxford Road
Manchester M13 9PL
Email: warren.mansell@manchester.ac.uk
Tel: +44 (0) 161 275 8589
Website: http://www.psych-sci.manchester.ac.uk/staff/131406
Advanced notice of a new transdiagnostic therapy manual, authored by Carey, Mansell & Tai - Principles-Based Counselling and Psychotherapy: A Method of Levels Approach
Available Now
Check www.pctweb.org for further information on Perceptual Control Theory
philip (8.31.2015)–
PY: …almost there…
…we need a closer look at the reference signal.
RM: This is a little more comprehensible to me, given my very meager understanding of biochemistry. I think the system above the dashed line is a lymphocyte; the stuff below the line is the environment of the lymphocyte (the blood?). The lymphocyte is controlling something about the virus. Since, as I understand it, lymphocytes eliminate viruses, or try to, the lymphocyte is controlling for getting whatever it perceives about the virus to zero. And it does it by producing antibodies. So what I see here is an attempt to model the control of viruses by lymphocytes and basing the structure of the model on what is known about the interactions between biochemical variables in the lymphocyte and its environment. Is that right?
Anyway, the diagrams are really nice, even if I don’t understand them. And you are to be commended for all the hard work you’ve put into this.
Best
Rick
–
Richard S. Marken
www.mindreadings.com
Author of Doing Research on Purpose.
Now available from Amazon or Barnes & Noble